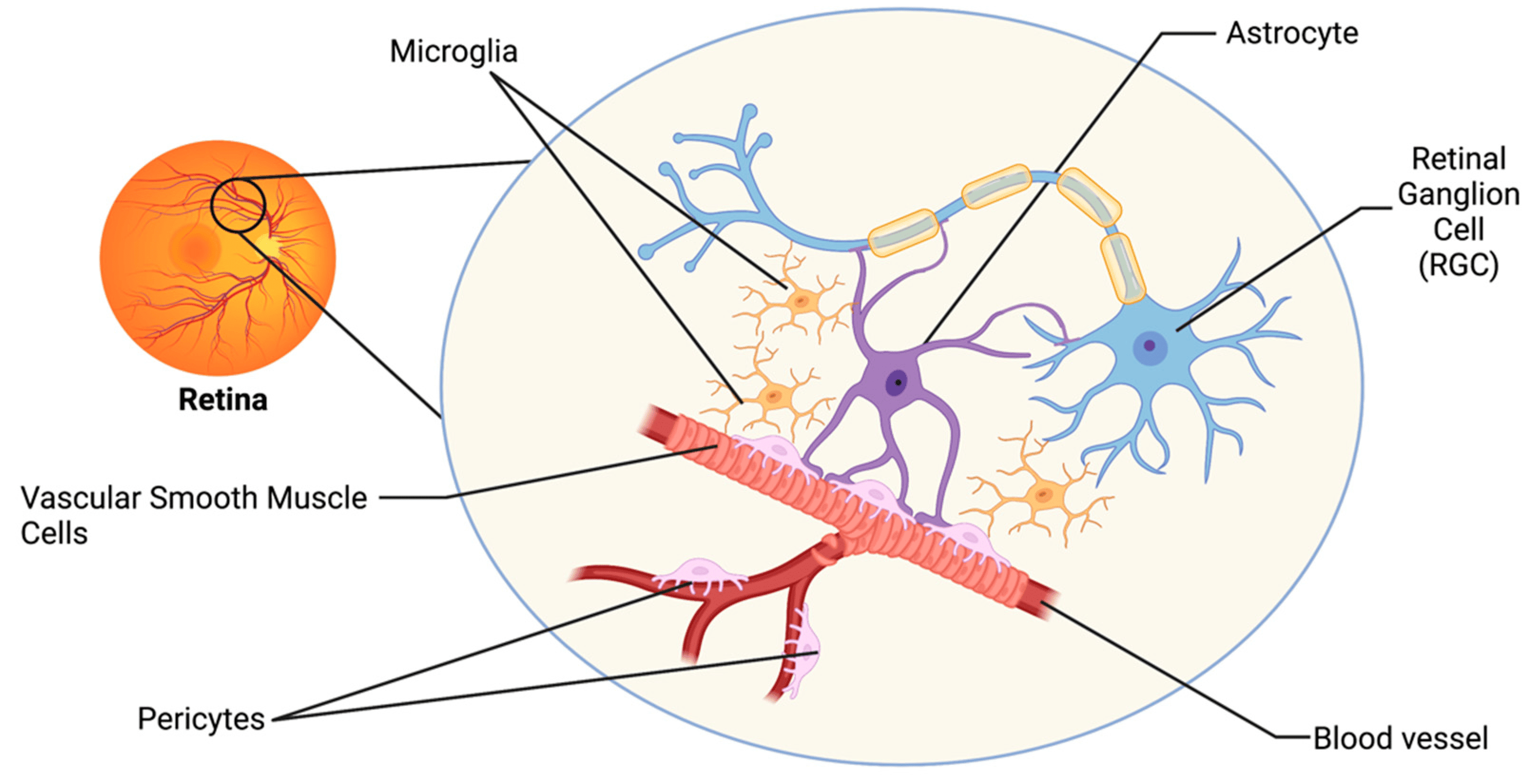
Astrocytes are the major glial cell type in the
non-myelinated optic nerve head (ONH) in most mammals and provide cellular support functions to
the axons while interfacing between connective tissue surfaces and surrounding
blood vessels.
Astrocytes form a mesh-like network on the
surface of the retina and tightly contact blood vessels and retinal ganglion cell (RGC) axons.
ONH astrocytes are responsible for the normal
maintenance of the extra-cellular matrix (ECM) in normal eyes. These cells are
sensitive to mechanical or ischemic factors and are important for the maintenance
of retinal ganglion physiology.
1A astrocytes provide structural support for
the axons, with type 1B cells providing a physiological interface between the
vitreous connective and vascular tissues.
In the normal ONH, astrocytes are considered to be quiescent.

In the lamina cribrosa, astrocytes form
lamellae, oriented perpendicular to the axons surrounding a core of fibroelastic
extracellular matrix.

Astrocytes supply energy substrate to axons in
the optic nerve and maintain extracellular pH and ion homeostasis in the
periaxonal space. Sodium channels in astrocytes participate in Na+ homeostasis,
providing a path for Na+ entry into the cytoplasm.
In astrocytes, voltage-gated calcium channels
deliver Ca2+ into the cytoplasm and participate in generation of glial Ca2+
signals.
Astrocytes maintain the scant periaxonal ECM
consisting of glycoproteins, such as laminin and proteoglycans.
Astrocytes express a wide variety of growth
factors and receptors, many of which serve as trophic and survival factors for
neurons.
Astrocytes are the cells responsible for many
pathological changes in the glaucomatous optic nerve head (ONH).
Astrocytes become reactive in response to
injury or disease and participate in the formation of a glial scar, which does
not support axonal survival or growth.
The major hallmarks of a reactive astrocyte are
an enlarged cell body and a thick network of processes with increased
expression of GFAP and vimentin.
Reactive astrocytes increase expression of
various cell surface molecules that play important roles in cell–cell recognition
and in cell adhesion to substrates, as well as various growth factors,
cytokines, and receptors. Reactive astrocytes express many new ECM proteins
such as laminin, tenascin C, and proteoglycans.
Reactive astrocytes in the glaucomatous ONH are
large rounded cells with many thick processes which expresses increased amounts
of GFAP, vimentin, and HSP27.
Recent evidence suggests that optic nerve head
astrocytes, which have long been recognized as important components of the
optic nerve head, may underlie this process and be central to the initiation of glaucomatous optic atrophy. These cells probably have a direct toxic effect on the RGC axons.
In glaucoma, reactive astrocytes have been
shown to migrate from the cribriform plates into the nerve bundles and
synthesize neurotoxic mediators such as nitric oxide (NO) and TNF-α, which may
be released near axons causing neuronal damage.
Reactive astrocytes in the ONH express large
amounts of elastin, leading to elastotic degeneration of the ECM in glaucoma
and loss of resiliency and deformability in response to elevated IOP.
ONH astrocytes may offer neuroprotection in the
optic nerve by releasing glutathione (GSH) and antioxidant enzymes to eliminate
the products of chronic oxidative stress that may be contributing to the
progression of neurodegeneration in POAG.
Astrocyte dysfunction could disrupt axoplasmic
transport and initiate the changes in cribrosal physiology that are said to be secondary
to the mechanical effects of raised IOP or to ischemic damage secondary to
optic disc hypoperfusion.
This hypothesis implies that significant disturbances of astrocyte metabolism may predispose to axon loss and initiate changes in cribrosal structure. Thus, the collapse of cribrosal beams, rather than initiating axon loss, may be as much the result of astrocyte fallout.







